
Page 227 - Hematology_ Basic Principles and Practice ( PDFDrive )
P. 227
Chapter 17 Control of Cell Division 179
ACP/C CDC20
Cyclin B
CDK1
SCF Cyclin F
Metaphase
Anaphase ACP/C CDH1
Prophase
Telophase
FOXM1
B–MYB Mitosis
Cyclin A
CDK1 Dream G 0
G 2 G 1
RB
S
Cyclin D
CDK4/6
E2F1–3
Cyclin A Cyclin E
CDK2 CDK2
SCF SKP2
SCF FBXW7
SCF betaTRCP
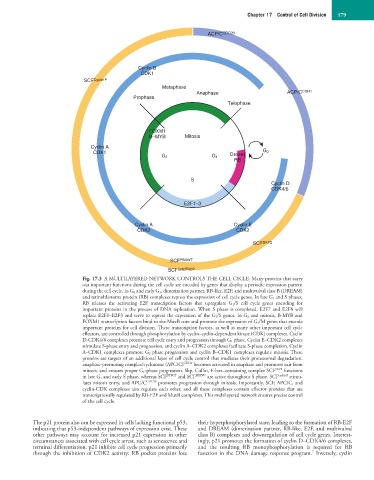
Fig. 17.3 A MULTILAYERED NETWORK CONTROLS THE CELL CYCLE. Many proteins that carry
out important functions during the cell cycle are encoded by genes that display a periodic expression pattern
during the cell cycle. In G 0 and early G 1, dimerization partner, RB-like, E2F, and multivulval class B (DREAM)
and retinoblastoma protein (RB) complexes repress the expression of cell cycle genes. In late G 1 and S phases,
RB releases the activating E2F transcription factors that upregulate G 1/S cell cycle genes encoding for
important proteins in the process of DNA replication. When S phase is completed, E2F7 and E2F8 will
replace E2F1–E2F3 and serve to repress the expression of the G 1/S genes. In G 2 and mitosis, B-MYB and
FOXM1 transcription factors bind to the MuvB core and promote the expression of G 2/M genes that encode
important proteins for cell division. These transcription factors, as well as many other important cell cycle
effectors, are controlled through phosphorylation by cyclin–cyclin-dependent kinase (CDK) complexes. Cyclin
D-CDK4/6 complexes promote cell cycle entry and progression through G 1 phase. Cyclin E–CDK2 complexes
stimulate S-phase entry and progression, and cyclin A–CDK2 complexes facilitate S-phase completion. Cyclin
A–CDK1 complexes promote G 2 phase progression and cyclin B–CDK1 complexes regulate mitosis. These
proteins are targets of an additional layer of cell cycle control that mediates their proteasomal degradation.
anaphase-promoting complex/cyclosome (APC/C) CDH1 becomes activated in anaphase and promotes exit from
mitosis, and ensures proper G 1-phase progression. Skp, Cullin, F-box–containing complex SCF SKP2 functions
in late G 1 and early S phase, whereas SCF βTRCP and SCF FBXW7 are active throughout S phase. SCF cyclin F stimu-
lates mitosis entry, and APC/C CDC20 promotes progression through mitosis. Importantly, SCF, APC/C, and
cyclin-CDK complexes also regulate each other, and all these complexes contain effector proteins that are
transcriptionally regulated by RB-E2F and MuvB complexes. This multilayered network ensures precise control
of the cell cycle.
The p21 protein also can be expressed in cells lacking functional p53, their hyperphosphorylated state, leading to the formation of RB-E2F
indicating that p53-independent pathways of expression exist. These and DREAM (dimerization partner, RB-like, E2F, and multivulval
other pathways may account for increased p21 expression in other class B) complexes and downregulation of cell cycle genes. Interest-
circumstances associated with cell cycle arrest, such as senescence and ingly, p21 promotes the formation of cyclin D–CDK4/6 complexes,
terminal differentiation. p21 inhibits cell cycle progression primarily and the resulting RB monophosphorylation is required for RB
7
through the inhibition of CDK2 activity: RB pocket proteins lose function in the DNA damage response program. Inversely, cyclin