
Page 126 - Hematology_ Basic Principles and Practice ( PDFDrive )
P. 126
Chapter 9 Hematopoietic Stem Cell Biology 97
ETV2
SCL Hemogenic
endothelium
Mesoderm
SCL/TAL1
LT-HSC LMO2
GATA2
–
+
c-Kit Lin Sca-1 Thy-1.1 FLK2 CD34 CD150 + RUNX1
–
–
+
lo
MLL
Multipotent c-Kit Lin Sca-1 Thy-1.1 FLK2 CD34 CD150 + ETV6
ST-HSC
BMI1
+
+
–
lo
–
+
GFI1
MPP
+
c-Kit Lin Sca-1 Thy-1.1 FLK2 CD34 CD150 –
–
+
+
+
–
CLP
–
lo
c-Kit Lin Sca-1 II7r +
lo
CMP
–
+
c-Kit Lin Sca-1 Fc R lo CD34 +
lo
IKAROS IKAROS
Oligopotent c-Kit Lin Sca-1 – – c-Kit Lin Sca-1 – + PAX5 NOTCH
PU.1
PU.1
MEP
GMP
E2A
–
+
+
–
EBF
TCF1
–
Fc R
+
Fc R
CD34
CD34
GATA3
GATA1 PU.1 PU.1 PU.1 PU.1 PU.1 BCL11A
GATA2 GATA2 C/EBP C/EBP (EVI9)
FOG1 GATA1
Committed precursors GATA1 GATA1 GATA1 C/EBP GFI1 XBP1
FOG1
GFI1B
EKLF GFI1B C/EBP
Mature cells RBC Megakaryocyte Mast Basophil Eosinophil Neutrophil Monocyte/ Dendritic B lymphocyte NK cell
T lymphocyte
cell
cell
Macrophage
FLI1
NFE2
Platelets
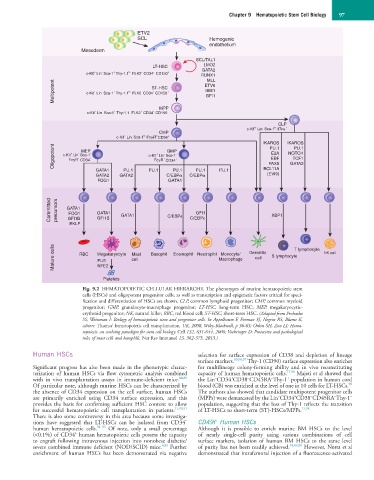
Fig. 9.2 HEMATOPOIETIC CELLULAR HIERARCHY. The phenotypes of murine hematopoietic stem
cells (HSCs) and oligopotent progenitor cells, as well as transcription and epigenetic factors critical for speci-
fication and differentiation of HSCs are shown. CLP, common lymphoid progenitor; CMP, common myeloid
progenitor; GMP, granulocyte-macrophage progenitor; LT-HSC, long-term HSC; MEP, megakaryocytic-
erythroid progenitor; NK, natural killer; RBC, red blood cell; ST-HSC, short-term HSC. (Adapted from Prohaska
SS, Weissman I: Biology of hematopoietic stem and progenitor cells. In Appelbaum F, Forman SJ, Negrin RS, Blume K,
editors: Thomas’ hematopoietic cell transplantation, UK, 2008, Wiley-Blackwell, p 36-63; Orkin SH, Zon LI: Hema-
topoiesis: an evolving paradigm for stem cell biology. Cell 132, 631-644, 2008; Voehringer D: Protective and pathological
roles of mast cells and basophils. Nat Rev Immunol 13, 362-375, 2013.)
Human HSCs selection for surface expression of CD38 and depletion of lineage
surface markers. 89,96,97 Thy-1 (CD90) surface expression also enriches
Significant progress has also been made in the phenotypic charac- for multilineage colony-forming ability and in vivo reconstituting
terization of human HSCs via flow cytometric analysis combined capacity of human hematopoietic cells. 17,98 Majeti et al showed that
+
−
−
−
+
with in vivo transplantation assays in immune-deficient mice. 88,89 the Lin CD34 CD38 CD45RA Thy-1 population in human cord
98
Of particular note, although murine HSCs can be characterized by blood (CB) was enriched at the level of one in 10 cells for LT-HSCs.
the absence of CD34 expression on the cell surface, human HSCs The authors also showed that candidate multipotent progenitor cells
−
−
−
+
−
are primarily enriched using CD34 surface expression, and this (MPPs) were demarcated by the Lin CD34 CD38 CD45RA Thy-1
provides the basis for confirming sufficient HSC content to allow population, suggesting that the loss of Thy-1 reflects the transition
for successful hematopoietic cell transplantation in patients. 17,90,91 of LT-HSCs to short-term (ST)-HSCs/MPPs. 17,98
There is also some controversy in this area because some investiga-
+
−
tions have suggested that LT-HSCs can be isolated from CD34 CD49f Human HSCs
human hematopoietic cells. 92–95 Of note, only a small percentage Although it is possible to enrich murine BM HSCs to the level
+
(<0.1%) of CD34 human hematopoietic cells possess the capacity of nearly single-cell purity using various combinations of cell
to engraft following intravenous injection into nonobese diabetic/ surface markers, isolation of human BM HSCs to the same level
severe combined immune deficient (NOD/SCID) mice. 4,89 Further of purity has not been readily achieved. 74,80,99 However, Notta et al
enrichment of human HSCs has been demonstrated via negative demonstrated that intrafemoral injection of a fluorescence-activated