
Page 267 - Hematology_ Basic Principles and Practice ( PDFDrive )
P. 267
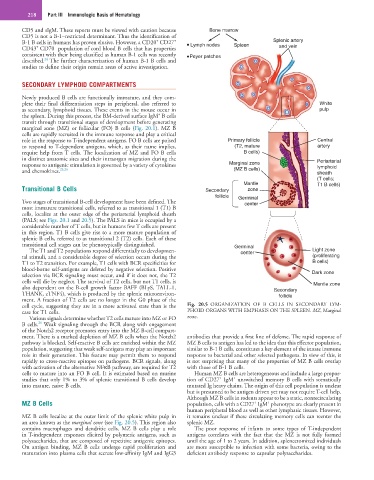
218 Part III Immunologic Basis of Hematology
CD5 and sIgM. These reports must be viewed with caution because Bone marrow
CD5 is not a B-1–restricted determinant. Thus the identification of
+
+
B-1 B cells in humans has proven elusive. However, a CD20 CD27 Lymph nodes Spleen Splenic artery
+
−
CD43 CD70 population of cord blood B cells that has properties and vein
consistent with their being classified as human B-1 cells was recently Peyer patches
24
described. The further characterization of human B-1 B cells and
studies to define their origin remain areas of active investigation.
SECONDARY LYMPHOID COMPARTMENTS
Newly produced B cells are functionally immature, and they com-
plete their final differentiation steps in peripheral, also referred to White
as secondary, lymphoid tissues. These events in the mouse occur in pulp
+
the spleen. During this process, the BM-derived surface IgM B cells
transit through transitional stages of development before generating
marginal zone (MZ) or follicular (FO) B cells (Fig. 20.1). MZ B
cells are rapidly recruited in the immune response and play a critical
role in the response to T-independent antigens. FO B cells are poised Primary follicle Central
to respond to T-dependent antigens, which, as their name implies, (T2, mature artery
require help from T cells. The localization of MZ and FO B cells B cells)
in distinct anatomic sites and their intraorgan migration during the Periarterial
response to antigenic stimulation is governed by a variety of cytokines Marginal zone lymphoid
and chemokines. 25,26 (MZ B cells) sheath
(T cells;
Mantle T1 B cells)
Transitional B Cells Secondary zone
follicle Germinal
Two stages of transitional B-cell development have been defined. The center
most immature transitional cells, referred to as transitional 1 (T1) B
cells, localize at the outer edge of the periarterial lymphoid sheath
(PALS; see Figs. 20.1 and 20.5). The PALS in mice is occupied by a
considerable number of T cells, but in humans few T cells are present
in this region. T1 B cells give rise to a more mature population of
splenic B cells, referred to as transitional 2 (T2) cells. Each of these
transitional cell stages can be phenotypically distinguished. Germinal
The T1 and T2 populations respond differentially to developmen- center Light zone
tal stimuli, and a considerable degree of selection occurs during the (proliferating
T1 to T2 transition. For example, T1 cells with BCR specificities for B cells)
blood-borne self-antigens are deleted by negative selection. Positive
selection via BCR signaling must occur, and if it does not, the T2 Dark zone
cells will die by neglect. The survival of T2 cells, but not T1 cells, is Mantle zone
also dependent on the B-cell growth factor BAFF (BLyS, TALL-1, Secondary
THANK, zTNF4), which is produced by the splenic microenviron- follicle
ment. A fraction of T2 cells are no longer in the G0 phase of the
cell cycle, suggesting they are in a more activated state than is the Fig. 20.5 ORGANIZATION OF B CELLS IN SECONDARY LYM-
case for T1 cells. PHOID ORGANS WITH EMPHASIS ON THE SPLEEN. MZ, Marginal
Various signals determine whether T2 cells mature into MZ or FO zone.
26
B cells. Weak signaling through the BCR along with engagement
of the Notch2 receptor promotes entry into the MZ B-cell compart-
ment. There is a marked depletion of MZ B cells when the Notch2 antibodies that provide a first line of defense. The rapid response of
pathway is blocked. Self-reactive B cells are enriched within the MZ MZ B cells to antigen has led to the idea that this effector population,
population, suggesting that weak self-antigens may play an important similar to B-1 B cells, constitutes a key element of the innate immune
role in their generation. This feature may permit them to respond response to bacterial and other selected pathogens. In view of this, it
rapidly to cross-reactive epitopes on pathogens. BCR signals, along is not surprising that many of the properties of MZ B cells overlap
with activation of the alternative NFκB pathway, are required for T2 with those of B-1 B cells.
cells to mature into an FO B cell. It is estimated based on murine Human MZ B cells are heterogeneous and include a large propor-
+
+
studies that only 1% to 3% of splenic transitional B cells develop tion of CD27 IgM unswitched memory B cells with somatically
into mature, naive B cells. mutated Ig heavy chains. The origin of this cell population is unclear
but is presumed to be antigen driven yet may not require T-cell help.
Although MZ B cells in rodents appear to be a static, nonrecirculating
MZ B Cells population, cells with a CD27 IgM phenotype are clearly present in
+
+
human peripheral blood as well as other lymphatic tissues. However,
MZ B cells localize at the outer limit of the splenic white pulp in it remains unclear if these circulating memory cells can reenter the
an area known as the marginal zone (see Fig. 20.5). This region also splenic MZ.
contains macrophages and dendritic cells. MZ B cells play a role The poor response of infants to some types of T-independent
in T-independent responses elicited by polymeric antigens, such as antigens correlates with the fact that the MZ is not fully formed
polysaccharides, that are composed of repetitive antigenic epitopes. until the age of 1 to 2 years. In addition, splenectomized individuals
On antigen binding, MZ B cells undergo rapid proliferation and are more susceptible to infection with some bacteria, owing to the
maturation into plasma cells that secrete low-affinity IgM and IgG3 deficient antibody response to capsular polysaccharides.