
Page 1882 - Williams Hematology ( PDFDrive )
P. 1882
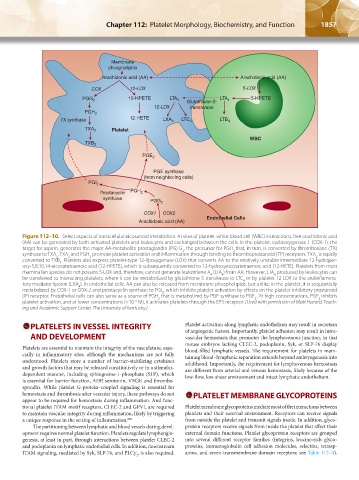
1856 Part XII: Hemostasis and Thrombosis Chapter 112: Platelet Morphology, Biochemistry, and Function 1857
Membrane
phospholipids
Arachidonic acid (AA) Arachidonic acid (AA)
COX 12-LOX 5-LOX
12-HPETE LTA LTA 5-HPETE
PGG 2 4 Glutathione-S- 4
12-LOX transferase
PGH 2
TX synthase 12-HETE LXA 4 LTC 4 LTB 4
TXA 2 Platelet
WBC
TXB 2
PGE 2
PGE synthase
(from neighboring cells)
PGI 2
Prostacyclin PGH 2
synthase
PGG 2
COX1 COX2
Endothelial Cells
Arachidonic acid (AA)
Figure 112–10. Select aspects of transcellular eicosanoid metabolism. At sites of platelet-white blood cell (WBC) interactions, free arachidonic acid
(AA) can be generated by both activated platelets and leukocytes and exchanged between the cells. In the platelet, cyclooxygenase 1 (COX-1), the
target for aspirin, generates the major AA-metabolite prostaglandin (PG) G , the precursor for PGH that, in turn, is converted by thromboxane (TX)
2
2
synthase to TXA . TXA and PGH promote platelet activation and inflammation through binding to thromboprostanoid (TP) receptors. TXA is rapidly
2
2
2
2
converted to TXB . Platelets also express platelet-type 12-lipoxygenase (LOX) that converts AA to the relatively unstable intermediate 12-hydroper-
2
oxy-5,8,10,14-eicosatetraenoic acid (12-HPETE), which is subsequently converted to 12-hydroxyeicosatetraenoic acid (12-HETE). Platelets from most
mammalian species do not possess 5-LOX and, therefore, cannot generate leukotriene A (LTA ) from AA. However, LTA produced by leukocytes can
4
4
4
be transferred to interacting platelets, where it can be metabolized by glutathione-S-transferase to LTC or by platelet 12-LOX to the antiinflamma-
4
tory mediator lipoxin (LXA ). In endothelial cells, AA can also be released from membrane phospholipids, but unlike in the platelet, it is sequentially
4
metabolized by COX-1 or COX-2 and prostacyclin synthase to PGI , which inhibits platelet activation by effects on the platelet inhibitory prostanoid
2
(IP) receptor. Endothelial cells can also serve as a source of PGH that is metabolized by PGE synthase to PGE . At high concentrations, PGE inhibits
2
2
2
platelet activation, and at lower concentrations (<10 M), it activates platelets through the EP3 receptor. (Used with permission of Matt Hazard, Teach-
–6
ing and Academic Support Center, The University of Kentucky.)
PLATELETS IN VESSEL INTEGRITY Platelet activation along lymphatic endothelium may result in secretion
of angiogenic factors. Importantly, platelet adhesion may result in intra-
AND DEVELOPMENT vascular hemostasis that promotes the lymphovenous junction, in that
mouse embryos lacking CLEC-2, podoplanin, Syk, or SLP-76 display
Platelets are essential to maintain the integrity of the vasculature, espe- blood-filled lymphatic vessels. The requirement for platelets in main-
cially in inflammatory sites, although the mechanisms are not fully taining blood–lymphatic separation extends beyond embryogenesis into
understood. Platelets store a number of barrier-stabilizing cytokines adulthood. Importantly, the requirement for lymphovenous hemostasis
and growth factors that may be released constitutively or in a stimulus- are different from arterial and venous hemostasis, likely because of the
dependent manner, including sphingosine-1-phosphate (S1P), which low-flow, low-shear environment and intact lymphatic endothelium.
is essential for barrier function, ADP, serotonin, VEGF, and thrombo-
spondin. While platelet G-protein–coupled signaling is essential for
hemostasis and thrombosis after vascular injury, these pathways do not PLATELET MEMBRANE GLYCOPROTEINS
appear to be required for hemostasis during inflammation. And func-
tional platelet ITAM motif receptors, CLEC-2 and GPVI, are required Platelet membrane glycoproteins mediate most of the interactions between
to maintain vascular integrity during inflammation, likely by triggering platelets and their external environment. Receptors can receive signals
a unique response in the setting of inflammation. 800 from outside the platelet and transmit signals inside. In addition, glyco-
The partitioning between lymphatic and blood vessels during devel- protein receptors receive signals from inside the platelet that affect their
opment requires normal platelet function. Platelets regulate lymphangio- external domain functions. Platelet glycoprotein receptors are grouped
genesis, at least in part, through interactions between platelet CLEC-2 into several different receptor families (integrins, leucine-rich glyco-
and podoplanin on lymphatic endothelial cells. In addition, downstream proteins, immunoglobulin cell adhesion molecules, selectins, tetrasp-
ITAM signaling, mediated by Syk, SLP-76, and PLCγ , is also required. anins, and seven-transmembrane domain receptors; see Table 112–4).
2
Kaushansky_chapter 112_p1829-1914.indd 1857 17/09/15 3:29 pm