
Page 1109 - Williams Hematology ( PDFDrive )
P. 1109
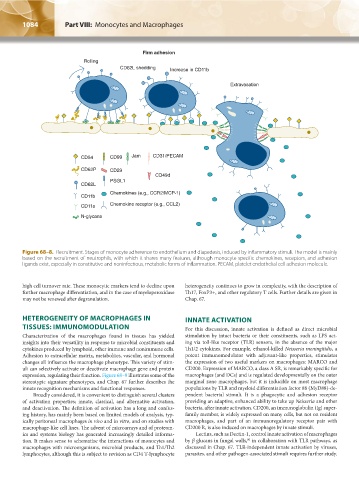
1084 Part VIII: Monocytes and Macrophages Chapter 68: Production, Distribution, and Activation of Monocytes and Macrophages 1085
Firm adhesion
Rolling
CD62L shedding Increase in CD11b
Extravasation
CD54 CD99 Jam CD31/PECAM
CD62P CD29
CD49d
PSGL1
CD62L
Chemokines (e.g., CCR2/MCP-1)
CD11b
CD11a Chemokine receptor (e.g., CCL2)
N-glycans
Figure 68–8. Recruitment. Stages of monocyte adherence to endothelium and diapedesis, induced by inflammatory stimuli. The model is mainly
based on the recruitment of neutrophils, with which it shares many features, although monocyte-specific chemokines, receptors, and adhesion
ligands exist, especially in constitutive and noninfectious, metabolic forms of inflammation. PECAM, platelet endothelial cell adhesion molecule.
high cell turnover rate. These monocytic markers tend to decline upon heterogeneity continues to grow in complexity, with the description of
further macrophage differentiation, and in the case of myeloperoxidase Th17, FoxP3+, and other regulatory T cells. Further details are given in
may not be renewed after degranulation. Chap. 67.
HETEROGENEITY OF MACROPHAGES IN INNATE ACTIVATION
TISSUES: IMMUNOMODULATION For this discussion, innate activation is defined as direct microbial
Characterization of the macrophages found in tissues has yielded stimulation by intact bacteria or their constituents, such as LPS act-
insights into their versatility in response to microbial constituents and ing via toll-like receptor (TLR) sensors, in the absence of the major
cytokines produced by lymphoid, other immune and nonimmune cells. Th1/2 cytokines. For example, ethanol-killed Neisseria meningitidis, a
Adhesion to extracellular matrix, metabolites, vascular, and hormonal potent immunomodulator with adjuvant-like properties, stimulates
changes all influence the macrophage phenotype. This variety of stim- the expression of two useful markers on macrophages: MARCO and
uli can selectively activate or deactivate macrophage gene and protein CD200. Expression of MARCO, a class A SR, is remarkably specific for
expression, regulating their function. Figure 68–9 illustrates some of the macrophages (and DCs) and is regulated developmentally on the outer
stereotypic signature phenotypes, and Chap. 67 further describes the marginal zone macrophages, but it is inducible on most macrophage
innate recognition mechanisms and functional responses. populations by TLR and myeloid differentiation factor 88 (MyD88)-de-
Broadly considered, it is convenient to distinguish several clusters pendent bacterial stimuli. It is a phagocytic and adhesion receptor
of activation properties; innate, classical, and alternative activation, providing an adaptive, enhanced ability to take up Neisseria and other
and deactivation. The definition of activation has a long and confus- bacteria, after innate activation. CD200, an immunoglobulin (Ig) super-
ing history, has mainly been based on limited models of analysis, typ- family member, is widely expressed on many cells, but not on resident
ically peritoneal macrophages in vivo and in vitro, and on studies with macrophages, and part of an immunoregulatory receptor pair with
macrophage-like cell lines. The advent of microarrays and of proteom- CD200 R, is also induced on macrophages by innate stimuli.
ics and systems biology has generated increasingly detailed informa- Lectins, such as Dectin-1, control innate activation of macrophages
80
tion. It makes sense to schematize the interactions of monocytes and by β glucans in fungal walls, in collaboration with TLR pathways, as
macrophages with microorganisms, microbial products, and Th1/Th2 discussed in Chap. 67. TLR-independent innate activation by viruses,
lymphocytes, although this is subject to revision as CD4 T-lymphocyte parasites, and other pathogen-associated stimuli requires further study.
Kaushansky_chapter 68_p1075-1088.indd 1084 9/17/15 3:42 PM