
Page 612 - Williams Hematology ( PDFDrive )
P. 612
586 Part VI: The Erythrocyte Chapter 41: Folate, Cobalamin, and Megaloblastic Anemias 587
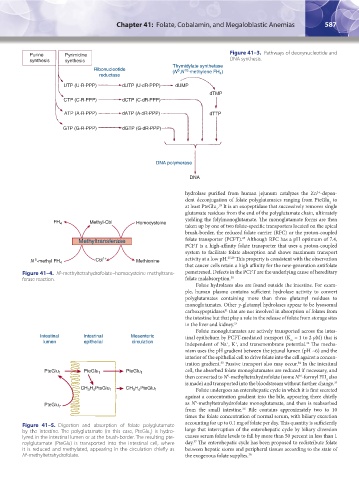
Purine Pyrimidine Figure 41–3. Pathways of deoxynucleotide and
synthesis synthesis DNA synthesis.
Thymidylate synthetase
Ribonucleotide (N ,N -methylene FH )
5
10
reductase 4
UTP (U·R·PPP) dUTP (U·dR·PPP) dUMP
dTMP
CTP (C·R·PPP) dCTP (C·dR·PPP)
ATP (A·R·PPP) dATP (A·dR·PPP) dTTP
GTP (G·R·PPP) dGTP (G·dR·PPP)
DNA polymerase
DNA
2+
hydrolase purified from human jejunum catalyzes the Zn -depen-
dent deconjugation of folate polyglutamates ranging from PteGlu to
2
28
at least PteGlu . It is an exopeptidase that successively removes single
7
glutamate residues from the end of the polyglutamate chain, ultimately
yielding the folylmonoglutamate. The monoglutamate forms are then
FH 4 Methyl-Cbl Homocysteine
taken up by one of two folate-specific transporters located on the apical
brush-border, the reduced folate carrier (RFC) or the proton-coupled
25
Methyltransferase folate transporter (PCFT). Although RFC has a pH optimum of 7.4,
PCFT is a high-affinity folate transporter that uses a proton-coupled
system to facilitate folate absorption and shows maximum transport
5
N -methyl FH 4 Cbl 1+ Methionine activity at a low pH. 25,29 This property is consistent with the observation
that cancer cells retain a high affinity for the new-generation antifolate
Figure 41–4. N -methyltetrahydrofolate–homocysteine methyltrans- pemetrexed. Defects in the PCFT are the underlying cause of hereditary
5
ferase reaction. folate malabsorption. 30
Folate hydrolases also are found outside the intestine. For exam-
ple, human plasma contains sufficient hydrolase activity to convert
polyglutamates containing more than three glutamyl residues to
monoglutamates. Other γ-glutamyl hydrolases appear to be lysosomal
carboxypeptidases that are not involved in absorption of folates from
31
the intestine but that play a role in the release of folate from storage sites
in the liver and kidney. 25
Folate monoglutamates are actively transported across the intes-
Intestinal Intestinal Mesenteric tinal epithelium by PCFT-mediated transport (K = 1 to 2 μM) that is
lumen epithelial circulation m
+
independent of Na , K , and transmembrane potential. The mecha-
32
+
nism uses the pH gradient between the jejunal lumen (pH ~6) and the
interior of the epithelial cell to drive folate into the cell against a concen-
34
33
tration gradient. Passive transport also may occur. In the intestinal
PteGlu 1 PteGlu 1 PteGlu 1 cell, the absorbed folate monoglutamates are reduced if necessary, and
10
then converted to N -methyltetrahydrofolate (some N -formyl FH also
5
4
is made) and transported into the bloodstream without further change. 35
CH H PteGlu 1 CH H PteGlu 1 Folate undergoes an enterohepatic cycle in which it is first secreted
3 4
3 4
against a concentration gradient into the bile, appearing there chiefly
5
PteGlu 7 as N -methyltetrahydrofolate monoglutamate, and then is reabsorbed
from the small intestine. Bile contains approximately two to 10
36
times the folate concentration of normal serum, with biliary excretion
Figure 41–5. Digestion and absorption of folate polyglutamate accounting for up to 0.1 mg of folate per day. This quantity is sufficiently
by the intestine. The polyglutamate (in this case, PteGlu ) is hydro- large that interruption of the enterohepatic cycle by biliary diversion
7
lyzed in the intestinal lumen or at the brush-border. The resulting pte- causes serum folate levels to fall by more than 50 percent in less than 1
37
roylglutamate (PteGlu) is transported into the intestinal cell, where day. The enterohepatic cycle has been proposed to redistribute folate
it is reduced and methylated, appearing in the circulation chiefly as between hepatic stores and peripheral tissues according to the state of
N -methyltetrahydrofolate. the exogenous folate supplies. 38
5
Kaushansky_chapter 41_p0583-0616.indd 587 9/17/15 6:23 PM