
Page 764 - Williams Hematology ( PDFDrive )
P. 764
738 Part VI: The Erythrocyte Chapter 48: The Thalassemias: Disorders of Globin Synthesis 739
Hgb Seal Rock Hgb Pakse ? Hgb A conditions, which can occur in any racial groups, include α-thalassemia
A(Glu) (Tyr) UUA(Leu) UGA associated with mental retardation or leukemia. Their importance lies
GAA UAU (terminate) with the diagnostic problems they may present and, more importantly,
UAG the light that elucidation of the α-thalassemia pathology may shed on
broader disease mechanisms.
HgbA Molecular Pathology of the α-Thalassemia Mental
UAA Retardation Syndrome
a142
The first descriptions of noninherited forms of α-thalassemia associ-
ated with mental retardation suggested the lesions involving the α-glo-
bin gene locus were acquired in the paternal germ cells and that their
AAA CAA UCA molecular pathology might help elucidate the associated developmental
(Lys) (Gln) (Ser) changes. Two separate syndromes of this type now are evident. In one
156
Hgb lcaria Hgb CS Hgb Koya Dora
group of patients, long deletions involve the α-globin gene cluster and
157
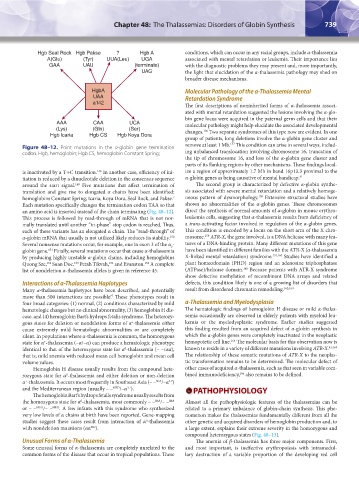
Figure 48–12. Point mutations in the α-globin gene termination remove at least 1 Mb. This condition can arise in several ways, includ-
codon. Hgb, hemoglobin; Hgb CS, hemoglobin Constant Spring; ing unbalanced translocation involving chromosome 16, truncation of
the tip of chromosome 16, and loss of the α-globin gene cluster and
parts of its flanking regions by other mechanisms. These findings local-
is inactivated by a T→C transition. In another case, efficiency of ini- ize a region of approximately 1.7 Mb in band 16p13.3 proximal to the
146
tiation is reduced by a dinucleotide deletion in the consensus sequence α-globin genes as being causative of mental handicap. 41
around the start signal. Five mutations that affect termination of The second group is characterized by defective α-globin synthe-
149
translation and give rise to elongated α chains have been identified: sis associated with severe mental retardation and a relatively homoge-
158
hemoglobins Constant Spring, Icaria, Koya Dora, Seal Rock, and Pakse. neous pattern of dysmorphology. Extensive structural studies have
7
Each mutation specifically changes the termination codon TAA so that shown no abnormalities of the α-globin genes. These chromosomes
an amino acid is inserted instead of the chain terminating (Fig. 48–12). direct the synthesis of normal amounts of α-globin in mouse erythro-
This process is followed by read-through of mRNA that is not nor- leukemia cells, suggesting that α-thalassemia results from deficiency of
mally translated until another “in-phase” stop codon is reached. Thus, a trans-activating factor involved in regulation of the α-globin genes.
each of these variants has an elongated α chain. The “read-through” of This condition is encoded by a locus on the short arm of the X chro-
159
α-globin mRNA that usually is not utilized likely reduces its stability. mosome. ATR-X, the gene involved, is a DNA helicase with many fea-
150
Several nonsense mutations occur, for example, one in exon 3 of the α - tures of a DNA-binding protein. Many different mutations of this gene
2
globin gene. Finally, several mutations occur that cause α-thalassemia have been identified in different families with the ATR-X (α-thalassemia
151
by producing highly unstable α-globin chains, including hemoglobins X-linked mental retardation) syndrome. 131,160 Studies have identified a
Quong Sze, Suan Doc, Petah Tikvah, and Evanston. A complete plant homeodomain (PHD) region and an adenosine triphosphatase
154
152
153
155
161
list of nondeletion α-thalassemia alleles is given in reference 45. (ATPase)/helicase domain. Because patients with ATR-X syndrome
show defective methylation of recombinant DNA arrays and related
Interactions of α-Thalassemia Haplotypes defects, this condition likely is one of a growing list of disorders that
Many α-thalassemia haplotypes have been described, and potentially result from disordered chromatin remodeling. 162,163
more than 500 interactions are possible! These phenotypes result in
7
four broad categories: (1) normal, (2) conditions characterized by mild α-Thalassemia and Myelodysplasia
hematologic changes but no clinical abnormality, (3) hemoglobin H dis- The hematologic findings of hemoglobin H disease or mild α-thalas-
ease, and (4) hemoglobin Bart’s hydrops fetalis syndrome. The heterozy- semia occasionally are observed in elderly patients with myeloid leu-
gous states for deletion or nondeletion forms of α -thalassemia either kemia or the myelodysplastic syndrome. Earlier studies suggested
+
cause extremely mild hematologic abnormalities or are completely this finding resulted from an acquired defect of α-globin synthesis in
silent. In populations where α-thalassemia is common, the homozygous which the α-globin genes were completely inactivated in the neoplastic
164
state for α -thalassemia (–α/–α) can produce a hematologic phenotype hemopoietic cell line. The molecular basis for this observation now is
+
identical to that of the heterozygous state for α -thalassemia (– –/αα), known to reside in a variety of different mutations involving ATR-X. 41,165
0
that is, mild anemia with reduced mean cell hemoglobin and mean cell The relationship of these somatic mutations of ATR-X to the neoplas-
volume values. tic transformation remains to be determined. The molecular defect of
Hemoglobin H disease usually results from the compound hete- other cases of acquired α-thalassemia, such as that seen in variable com-
166
rozygous state for α -thalassemia and either deletion or non-deletion bined immunodeficiency, also remains to be defined.
0
α -thalassemia. It occurs most frequently in Southeast Asia (– – /–α )
+
SEA
3.7
and the Mediterranean region (usually – – MED /–α ). PATHOPHYSIOLOGY
3.7
The hemoglobin Bart’s hydrops fetalis syndrome usually results from
the homozygous state for α -thalassemia, most commonly – – SEA /– – SEA Almost all the pathophysiologic features of the thalassemias can be
0
or – – MED /– – MED . A few infants with this syndrome who synthesized related to a primary imbalance of globin-chain synthesis. This phe-
very low levels of α chains at birth have been reported. Gene-mapping nomenon makes the thalassemias fundamentally different from all the
studies suggest these cases result from interaction of α -thalassemia other genetic and acquired disorders of hemoglobin production and, to
0
with nondeletion mutations (αα ). a large extent, explains their extreme severity in the homozygous and
ND
compound heterozygous states (Fig. 48–13).
Unusual Forms of α-Thalassemia The anemia of β-thalassemia has three major components. First,
Some unusual forms of α-thalassemia are completely unrelated to the and most important, is ineffective erythropoiesis with intramedul-
common forms of the disease that occur in tropical populations. These lary destruction of a variable proportion of the developing red cell
Kaushansky_chapter 48_p0725-0758.indd 739 9/18/15 2:57 PM