
Page 133 - Clinical Immunology_ Principles and Practice ( PDFDrive )
P. 133
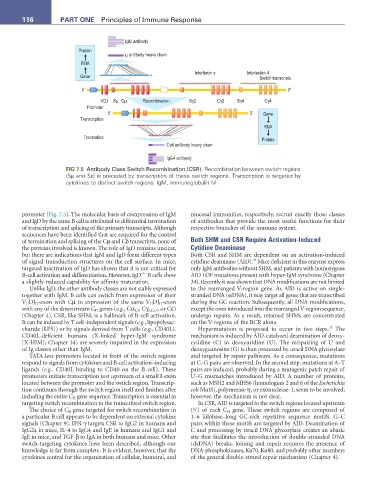
116 ParT ONE Principles of Immune Response
IgM antibody
Protein
µ antibody heavy chain
RNA
Interferon-γ Interleukin-4
Gene Switch transcripts
5' 3'
VDJ Sµ Cµ Recombination Sγ2 Cγ2 Sγ4 Cγ4
Promoter
5' 3' Gene
Transcription
RNA
Translation Protein
Cγ4 antibody heavy chain
IgG4 antibody
FIG 7.5 Antibody Class-Switch Recombination (CSR). Recombination between switch regions
(Sµ and Sε) is preceded by transcription of these switch regions. Transcription is targeted by
cytokines to distinct switch regions. IgM, Immunoglobulin M.
promoter (Fig. 7.5). The molecular basis of coexpression of IgM mucosal immunities, respectively, recruit exactly those classes
and IgD by the same B cell is attributed to differential termination of antibodies that provide the most useful functions for their
of transcription and splicing of the primary transcripts. Although respective branches of the immune system.
sequences have been identified that are required for the control
of termination and splicing of the Cµ and Cδ transcripts, none of Both SHM and CSR Require Activation-Induced
the proteins involved is known. The role of IgD remains unclear, Cytidine Deaminase
but there are indications that IgM and IgD form different types Both CSR and SHM are dependent on an activation-induced
28
of signal transduction structures on the cell surface. In mice, cytidine deaminase (AID). Mice deficient in this enzyme express
targeted inactivation of IgD has shown that it is not critical for only IgM antibodies without SHM, and patients with homozygous
−/−
B-cell activation and differentiation. However, IgD B cells show AID LOF mutations present with hyper-IgM syndrome (Chapter
a slightly reduced capability for affinity maturation. 34). Recently it was shown that DNA modifications are not limited
Unlike IgD, the other antibody classes are not stably expressed to the rearranged V-region gene. As AID is active on single-
together with IgM. B cells can switch from expression of their stranded DNA (ssDNA), it may target all genes that are transcribed
V H DJ H -exon with Cµ to expression of the same V H DJ H -exon during the GC reaction. Subsequently, all DNA modifications,
with any of the downstream C H genes (e.g., Cα 1,2 , Cγ 1,2,3,4 , or Cε) except the ones introduced into the rearranged V-region sequence,
(Chapter 4). CSR, like SHM, is a hallmark of B-cell activation. undergo repairs. As a result, retained SHMs are concentrated
It can be induced by T cell–independent signals (e.g., lipopolysac- on the V-regions of the BCR alone.
28
charide [LPS]) or by signals derived from T cells (e.g., CD40L). Hypermutation is proposed to occur in two steps. The
CD40L-deficient humans (X-linked hyper-IgM syndrome mechanism is induced by AID-catalyzed deamination of deoxy-
[X-HIM]; Chapter 34) are severely impaired in the expression cytidine (C) to deoxyuridine (U). The mispairing of U and
of Ig classes other than IgM. deoxyguanosine (G) is than processed by uracil DNA glycosylase
TATA-less promoters located in front of the switch regions and targeted by repair pathways. As a consequence, mutations
respond to signals from cytokines and B-cell activation–inducing at C–G pairs are observed. In the second step, mutations at A–T
ligands (e.g., CD40L binding to CD40 on the B cell). These pairs are induced, probably during a mutagenic patch repair of
promoters initiate transcription just upstream of a small Ι-exon U–G mismatches introduced by AID. A number of proteins,
located between the promoter and the switch region. Transcrip- such as MSH2 and MHS6 (homologues 2 and 6 of the Escherichia
tion continues through the switch region itself and finishes after coli MutS), polymerase η, or exonuclease-1, seem to be involved;
including the entire C H gene sequence. Transcription is essential in however, the mechanism is not clear.
targeting switch recombination to the transcribed switch region. In CSR, AID is targeted to the switch regions located upstream
The choice of C H gene targeted for switch recombination in (5’) of each C H gene. These switch regions are composed of
a particular B cell appears to be dependent on external cytokine 1–6 kilobase–long GC-rich repetitive sequence motifs. G–C
signals (Chapter 9). IFN-γ targets CSR to IgG2 in humans and pairs within these motifs are targeted by AID. Deamination of
IgG2a in mice, IL-4 to IgG4 and IgE in humans and IgG1 and C and processing by uracil DNA glycosylase creates an abasic
IgE in mice, and TGF-β to IgA in both humans and mice. Other site that facilitates the introduction of double-stranded DNA
switch-targeting cytokines have been described, although our (dsDNA) breaks. Joining and repair requires the presence of
knowledge is far from complete. It is evident, however, that the DNA-phosphokinases, Ku70, Ku80, and probably other members
cytokines central for the organization of cellular, humoral, and of the general double-strand repair mechanism (Chapter 4).