
Page 1091 - Williams Hematology ( PDFDrive )
P. 1091
1066 Part VIII: Monocytes and Macrophages Chapter 67: Structure, Receptors, and Functions of Monocytes and Macrophages 1067
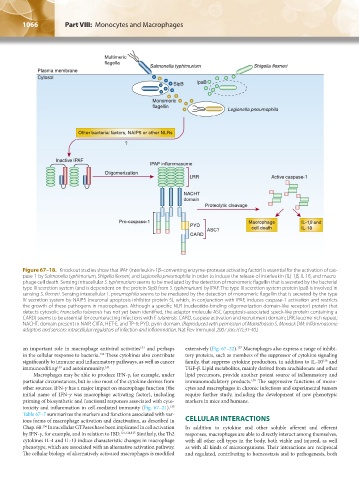
Figure 67–18. Knockout studies show that IPAF (interleukin-1β–converting enzyme-protease activating factor) is essential for the activation of cas-
pase-1 by Salmonella typhimurium, Shigella flexneri, and Legionella pneumophila in order to induce the release of interleukin (IL)-1β, IL-18, and macro-
phage cell death. Sensing intracellular S. typhimurium seems to be mediated by the detection of monomeric flagellin that is secreted by the bacterial
type III secretion system (and is dependent on the protein SipB from S. typhimurium) by IPAF. The type III secretion system protein IpaB is involved in
sensing S. flexneri. Sensing intracellular L. pneumophila seems to be mediated by the detection of monomeric flagellin that is secreted by the type
IV secretion system by NAIP5 (neuronal apoptosis inhibitor protein 5), which, in conjunction with IPAF, induces caspase-1 activation and restricts
the growth of these pathogens in macrophages. Although a specific NLR (nucleotide-binding oligomerization domain-like receptor) protein that
detects cytosolic Francisella tularensis has not yet been identified, the adaptor molecule ASC (apoptosis-associated speck-like protein containing a
CARD) seems to be essential for counteracting infections with F. tularensis. CARD, caspase activation and recruitment domain; LRR, leucine-rich repeat;
NACHT, domain present in NAIP, CIITA, HET-E, and TP-1; PYD, pyrin domain. (Reproduced with permission of Mariathasan S, Monack DM: Inflammasome
adaptors and sensors: intracellular regulators of infection and inflammation. Nat Rev Immunol 2007 Jan;7(1):31-40.)
an important role in macrophage antiviral activities and perhaps extensively (Fig. 67–22). Macrophages also express a range of inhibi-
131
137
in the cellular response to bacteria. These cytokines also contribute tory proteins, such as members of the suppressor of cytokine signaling
132
significantly to immune and inflammatory pathways, as well as cancer family, that suppress cytokine production, in addition to IL-10 and
138
immunoediting and autoimmunity. 134 TGF-β. Lipid metabolites, mainly derived from arachidonate and other
133
Macrophages may be able to produce IFN-γ, for example, under lipid precursors, provide another potent source of inflammatory and
particular circumstances, but in vivo most of the cytokine derives from immunomodulatory products. The suppressive functions of mono-
139
other sources. IFN-γ has a major impact on macrophage function (the cytes and macrophages in chronic infections and experimental tumors
initial name of IFN-γ was macrophage activating factor), including require further study, including the development of new phenotypic
priming of biosynthetic and functional responses associated with cyto- markers in mice and humans.
toxicity and inflammation in cell-mediated immunity (Fig. 67–21).
135
Table 67–7 summarizes the markers and functions associated with var-
ious forms of macrophage activation and deactivation, as described in CELLULAR INTERACTIONS
Chap. 68. Intracellular GTPases have been implicated in cell activation In addition to cytokine and other soluble afferent and efferent
136
by IFN-γ, for example, and in relation to IBD. 121,124,125 Similarly, the Th2 responses, macrophages are able to directly interact among themselves,
cytokines IL-4 and IL-13 induce characteristic changes in macrophage with all other cell types in the body, both viable and injured, as well
phenotype, which are associated with an alternative activation pathway. as with all kinds of microorganisms. Their interactions are reciprocal
The cellular biology of alternatively activated macrophages is modified and regulated, contributing to homeostasis and to pathogenesis, both
Kaushansky_chapter 67_p1043-1074.indd 1066 9/21/15 10:43 AM